1) нижнюю, к которой теперь и относится термин «А. з.», — с однообразной фауной и низкими температурами и
2) верхнюю, называемую теперь батиальной (или архибентальной), — с более разнообразной и богатой фауной и сравнительно высокими температурами (рис. 1).
Верхняя граница А. з. проходит большей частью на глубине 2000 м, в ряде районов — на глубинах от 1000 до 3000 м в соответствии с положением изотермы 4° С, которая разграничивает распределение эндемических форм. В полярных районах различие в фауне между А. з. и батиальной зоной нечеткое. В результате недавних исследований в глубоководных морях была выделена еще одна зона— ультраабиссальная, находящаяся на глубине более 6000— 7000 м (желоба и впадины), с присущей только ей фауной (см. Хадальная зона).1
А.
з. — самая большая экологическая единица
в мире — занимает 3/4
площади Мирового океана и немного более
половины всей площади Земли (см. табл.
1).

Условия
существования в А. з.
Факторы окружающей
среды. Условия существования в А. з.
очень однообразны, за исключением
источников питательных веществ.
Температура в А. з. колеблется от 0 до 2°
С; в каждом районе она постоянная, и
смена времен года на нее не влияет.
Низкая температура обусловливается
холодными водными массами, опускающимися
в полярных районах и распространяющимися вдоль дна к экватору. В частично закрытых глубоководных бассейнах, таких, как моря Средиземное, Японское и Суду, наблюдаются более высокие температуры воды у дна и настоящая абиссальная фауна отсутствует.
Соленость в А. з. равна 34,8±0,2%0. Щелочность, содержание фосфатов, двуокиси углерода и силикатов изменяются незначительно. Содержание кислорода зависит от притока насыщенных кислородом водных масс и составляет в среднем 5—6 см3/л в Атлантическом океане и 3,5—
4 см3/л в Тихом океане. У самого дна содержание кисло
рода может повышаться, что, вероятно, связано с биоло
гическими процессами. Давление в А. з. изменяется в
пределах 200 — 700 ат. В какой степени это влияет на
жизнь животных, пока еще не выяснено.
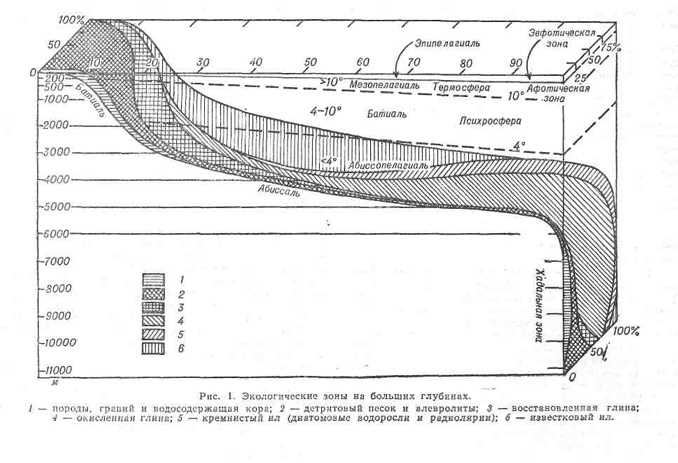
Субстрат. А. з. обычно является спокойной средой, где могут оседать очень мелкие частицы. Дно океана, если не считать подводных гор и горных хребтов, в основном покрыто слоем рыхлого глинистого ила довольно большой мощности, возможно, порядка сотен метров. Вблизи суши осадки состоят из терригенного и органического материалов, а также из остатков планктонных организмов. Дно более отдаленных участков океана покрыто в основном пелагическими осадками; в наибольшем удалении от суши осадки оседают очень медленно — менее 1 см/1000 лет. Переход от терригенных осадков к пелагическим происходит примерно на глубине 2000 м, что совпадает с верхней границей А. з.
На глубине около 4000 м (в средних и низких широтах) осадки обычно состоят из серого известкового ила, содержащего в большом количестве скелетные части пелагических фораминифер (например, глобигерин, Globigerina) и жгутиковых (особенно кок-колиты). Глобигериновые илы покрывают большую часть дна Атлантического и Индийского океанов, а также встречаются в южной части Тихого океана. На некоторых участках океанического дна в известковом иле преобладают оболочки птеропод. На большей части дна Тихого океана на глубинах более 4000 м известковые осадки в значительной степени растворились (см. Уровень карбонатной компенсации) и представляют собой коричневатую абиссальную глину (красную глину), состоящую в основном из алюмосиликата (рис. 2).
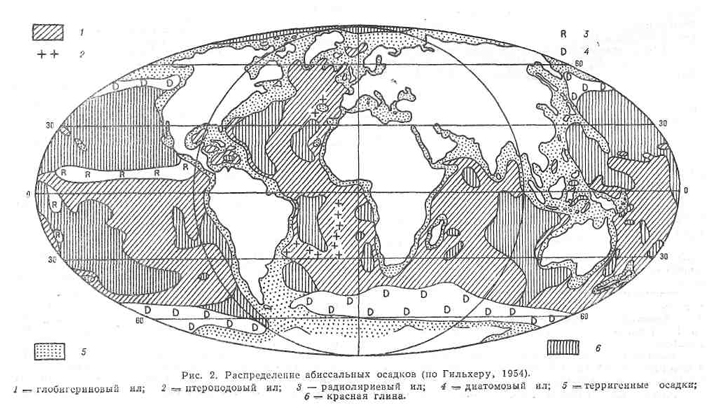
В более глубоководных районах Тихого и Индийского океанов осадки состоят в основном из кремниевых оболочек радиолярий. В Южном океане и северной части Тихого океана осадки в основном состоят из кремнистого ила, образовавшегося в результате накопления створок диатомей, живущих в холодных поверхностных водах. Скорость придонных вод небольшая — около 5 см/с, но на глубине около 3000 м и вокруг подводных гор скорость придонных течений может иногда увеличи ваться до 16 см/с, о чем свидетельствуют следы ряби на фотографиях дна глубоководных участков (см. Придонные океанические течения).
Источники питательных веществ. Питательные вещества для поддержания жизни в глубоководных районах поступают с суши и из верхних слоев океана, где они образуются в основном в результате жизнедеятельности фотоавтотрофного фитопланктона, что является начальным звеном цепи питания в океанах. Погружающиеся эпипелагические организмы потребляются бати-пелагической фауной. Однако остатки растительных и животных организмов и фекалий, погружающиеся на глубину более 2000 м, на пути ко дну потребляются незначительно, так как пелагическая жизнь в Д. з- очень бедна. Поэтому общее положение — на большие глубины поступает мало питательных веществ — справедливо лишь для глубоководных участков, находящихся на большом, расстоянии от суши или от районов с большой продуктивностью на поверхности.
Абиссальные гетеротрофные бактерии ассимилируют органические вещества и, вероятно, образуют главный источник питания или непосредственно, или в качестве второго звена новой цепи питания. Терри-генные осадки, содержащие остатки более богатой фауны, чем эвпелагические, могут быть вынесены далеко в океан мутьевыми потоками. В осадках некоторых А. з. тропических районов в изобилии находятся растительные осадки, которые, являясь источником питания, вызывают в А. з. концентрацию живых организмов. Затонувшие мертвые тела крупных животных, таких, например, как киты, создают временный местный источник питания, и этим можно отчасти объяснить неоднородное распределение многих абиссальных видов организмов.
Фауна.
Морфологические характеристики.
Животные А. з. обитают в темноте, в
спокойной среде, на мягком грунте, имеют
одинаковую окраску, чаще всего сероватую
или черную, в большинстве случаев они
хрупкие. У подвижных животных длинные
конечности. Сидячие медленно
переползают (что, возможно, определяется
их стремлением к перемещению в вышележащие
слои из придонного слоя, бедного
кислородом). Многие рыбы и ракообразные
слепы (или с очень развитыми глазами.
— Ред.); у некоторых рыб брюшные
плавники являются органами осязания.
Состав. Для А. з. характерны все основные пелагические виды беспозвоночных и некоторые бентосные виды рыб:
губки: Tetractinellida, Hexactinellida (Euplectellidae, Hyalonematidae);
кишечнополостные: Hydrozoa (Branchiocerianthus); Scyphozoa (Coronatae: Stephanoscyphus); Anthozoa (Gorgo-naria: Clavularidae; Pennatularia: Umbellundae); Actinia-ria; Madreporaria (одиночные виды); Antipataria (Bathy-pathes);
многощетинковые черви: Aphroditidae; Eunicidae;. Maldanidae; Terebellidae;
Echiuridae: Sipunculoidea; Priapuloidea; Nematoda; Nemertini, Bryozoa; Brachiopoda; Pogonophoa; Enterop-neusta; Pycnogonida;
ракообразные: Cirripedia (Scalpellum); Cumacea; Tanaidacea; Isopoda (Haplonlscus, Storthyngura, Arcturus); Amphipoda; Decapoda (Eryonidea, Paguridae, Galatheidea, Brachyura);
иглокожие: Crinoidea (стебельчатые формы); Asteroidea (Brisingidae, Pterasteridae, Porcellanasteridae); Ophiuroi-dea; Echinoidea (Echinothuridae, Spatangoidea); Hooth-rioidea (Elasipoda: Elpidia, Peniagone, Psychropotes, Bent-hodytes);
моллюски: Monoplacophora (Neopilina); Bivalvia (Nuculidae, Limopsidae, Cuspidariidae, Xylophaginidae); Gastropoda (Pleutotomella); Cephalopoda;
оболочники: Ascidiacea (Cukolus, Styela, Octacnetnus);
позвоночные: Selachii; Holocephali; Teleostei (Bathyp-teroidae, Synaphobranchidae, Macrouridae, Brotulidae, Liparidae, Ceratiidae).
Инфауна и эпифауна. Абиссальная фауна, подобно фауне холодных вод и инфауне мелководных участков, состоит из сравнительно немногих, но широко распространенных видов при относительно большом количестве особей внутри видов. Кроме того, имеется много видов, особенно в эпифауне, с пятнистым распределением. Эпифауна концентрируется на подводных горах и около них, где сильные придонные течения могут обнажать твердую поверхность или более грубозернистый субстрат. С помощью фотографий дна, сделанных в глубоководных районах, удалось установить, что, например, на одной подводной горе могут преобладать криноиды, а на другой — губки.
Типы питания. С увеличением глубины увеличивается количество потребителей суспензий (питающихся суспензионными наносами) и потребителей отложений (питающихся осевшими органическими веществами) относительно количества хищников и организмов, питающихся падалью. Porcellanasteridae, семейство илоедов, а также хищные морские звезды неизвестны на глубине меньше 1000 м. Они составляют четвертую часть видов морских звезд, которые обитают на глубине более 4000 м, и половину тех, которые обитают на глубине более 6000 м. Преобладают над всеми животными в А. з. голотурии-илоеды, Elasipoda. Некоторые абиссальные рыбы тоже питаются отложениями.
Плотность населения. Плотность населения А. з. очень низкая по сравнению с плотностью населения умеренных глубин. Выраженная в биомассе (количество вещества живых организмов в граммах на квадратный метр) плотность населения даже в наиболее плодородных абиссальных районах (в Южном океане и в северной части бореальной области) составляет не более 1% плотности населения плодородных прибрежных районов. На больших глубинах продуктивность может быть очень низкой (табл. 2).

Размножение. Скорость размножения животных организмов в А. з. кажется медленной; предполагается, что животные А. з. живут довольно долго. Пока еще неизвестно, относятся ли они к животным с обычным сезонным ритмом жизнедеятельности, но имеются указания на то, что для подлинно абиссальных видов это неважно, размножение может происходить в любое время года, причем одновременно созревает небольшое количество яиц. Для абиссальных видов обычным кажется непелагическое развитие, хотя можно ожидать, что личинки некоторых видов животных на определенной стадии развития могут свободно плавать в течение некоторого времени. Некоторые виды имеют пелагические личинки, а из бенто-абиссопелагических рыб некоторые виды, например, Synaphobranchidae и Ceratiidae, имеют личинки, живущие в верхней пелагической зоне.
Подразделения
Абиссальной зоны
Вертикальное подразделение. А. з. может быть разделена на верхнюю и нижнюю подзоны с границей на глубине около 4500 м, где происходит изменение видового состава. Ряд встречающихся во всем мире видов принадлежит исключительно нижней абиссальной подзоне. На глубине около 4500 м вода оказывается в недостаточной степени насыщенной кальцитом, и это может частично объяснить изменения в фауне.
Региональное распределение и подразделения. Распределение многих эндемических форм в А. з. объясняется однородными условиями среды и отсутствием определенных топографических барьеров; бентосные виды рыб, Anthozoa, иглокожие и оболочники имеют почти космополитическое распределение, в то время как ракообразные и моллюски — ограниченное. Наибольшая разница в составе фауны А. з. наблюдается по разным сторонам Американского материка (этот материк также представляет собой наиболее заметный барьер в А. з.) В отношении иглокожих другим барьером в распределении можно, по-видимому, считать глубоководные районы центральной части Тихого океана, имеющие очень скудные источники питания. Очень холодные глубинные воды (<0o С) в Южном океане, вероятно, являются причиной особого состава существующей там абиссальной фауны. Таким образом, можно отметить три основных зоо-географических подразделения А. з.:
-
Атлантический и Индийский океаны, включая юго-западную часть Тихого океана;
-
восточная часть Тихого океана;
-
антарктические глубоководные районы.
Более подробное зоогеографическое деление большей частью отражает зоогеографию батиальной зоны.
Второстепенные виды А. з. Довольно значительную часть фауны А. з. составляют многочисленные виды, проникающие из батиальной зоны. Такие виды были названы второстепенными абиссальными видами, или пришельцами, так как они хотя и живут в условиях среды А. з., но не могут там постоянно размножаться. Вероятно также, что распространению многих видов в глубоководные районы океана мешает ограниченное количество пищи.
Происхождение фауны. Эндемические формы. Только немногие группы более высоких таксономических форм обитают исключительно в глубоководных районах. Семейство актиний, как известно, обитает только на ультраабиссальных (хадальных) глубинах (см. Хадальная зона). Морские звезды, Porcellanasteridae, живут почти исключительно в А. з. (1000—7600 м). Две трети видов голотурий Elasipoda относятся к А. з., а одна треть — к батиальной. Моллюски класса Monoplacophora (Neopi-lina) исключительно батиально-абиссальные и представляют единственную абиссальную группу, прослеживающуюся с палеозоя. Доминирующие абиссальные группы, такие, как Elasipoda, в ископаемом состоянии не встречаются, и лишь немногие из других представленных семейств могут быть прослежены только с мезозоя.
Неэндемические формы. Принято считать, что низкая температура в современных А. з. в настоящее время является характерной особенностью, появившейся лишь в недавнее геологическое время, и что во время позднего мезозоя и раннетретичного периода температура в А. з. была около 10° С. Постепенное охлаждение в поздне-третичный период, достигшее максимума во время четвертичного оледенения, могло, очевидно, очень избирательно повлиять на фауну. Стенотермные абиссальные формы, приспособившиеся к жизни при температуре примерно 10° С, очевидно, при понижении температуры вымирали, хотя, может быть, они могли бы выжить в батиальной зоне. Таксономическая характеристика родов и видов иглокожих, многощетинковых червей и ракообразных8 встречав-
шихся на различных глубинах, привела к мысли, что они становились более приспособленными с увеличением глубины. Возможно, что А. з. со времени позднего мезозоя или раннетретичного периода постепенно населялась обитателями батиальных глубин. Эндемические классы и семейства глубоководных районов, например Hexactinel-lida, современные Crinoidea, Cidaroidea и Elasipoda, составляют, вероятно, наидревнейшие виды. Возможно, Neopillina тоже появились в А. з. не раньше раннетретичного периода. Другие группы, Gorgonaria, Astropectinida и макруриды, родственные мелководные формы которых живут в тропических прибрежных водах, возможно, тоже проникли в А. з. в раннетретичном периоде, прежде чем начало сказываться понижение температуры. Возможность переселения в позднетретичном периоде допускается для групп, широко распространенных в А. з. и имеющих родственные мелководные формы в более холодных районах. Должно быть и в позднечетвертичном (послеледниковом) периоде наблюдалось продолжающееся до сих пор проникновение тех видов, которые встречаются больше всего в полярных районах и лишь в ограниченном количестве в соседних глубоководных морях.
Ф. ДЖ, МЭДСЕН
Прим. ред.1 Значительный вклад в изучение А. з. Мирового океана сделан советскими исследователями. В частности, в 1949 г. экспедиция на «Витязе» установила наличие жизни на глубине 8000 м и тем самым опровергла существовавшие представления о невозможности жизни на глубинах более 6500 м (см. Беляев Г. М. Донная фауна наибольших глубин (ультраабиссали) Мирового океана. М., «Наука», 1966; Зенкевич Л. А. Биология морей СССР. М., Изд-во АН СССР, 1963; Иванов А. В. Материалы по экологии и географическому распространению погонофор. — Труды ИОАН СССР, 1960, т. 34, с. 3—20).
Источник: Океанографическая энциклопедия, 1974 год, с. 5-8
(прим. будьте бдительны - наука могла уйти далеко вперед)
Другие статьи из энциклопедии можно найти здесь.
В том числе - о литоральной зоне и типах осадков